Биосинтез белка в клетке кратко и понятно
Содержание:
Обмен веществ
Обмен веществ — важнейшее свойство живых организмов. Совокупность реакций обмена веществ, протекающих в организме, называется метаболизмом. Метаболизм состоит из реакций ассимиляции (пластического обмена, анаболизма) и реакций диссимиляции (энергетического обмена, катаболизма). Ассимиляция — совокупность реакций биосинтеза, протекающих в клетке, диссимиляция — совокупность реакций распада и окисления высокомолекулярных веществ, идущих с выделением энергии. Эти группы реакций взаимосвязаны: реакции биосинтеза невозможны без энергии, которая выделяется в реакциях энергетического обмена, реакции диссимиляции не идут без ферментов, образующихся в реакциях пластического обмена.
По типу обмена веществ организмы подразделяются на две группы: автотрофы и гетеротрофы. Автотрофы — организмы, способные синтезировать органические вещества из неорганических и использующие для этого синтеза или солнечную энергию, или энергию, выделяющуюся при окислении неорганических веществ. Гетеротрофы — организмы, использующие для своей жизнедеятельности органические вещества, синтезированные другими организмами. В качестве источника углерода автотрофы используют неорганические вещества (СО2), а гетеротрофы — экзогенные органические. Источники энергии: у автотрофов — энергия солнечного света (фотоавтотрофы) или энергия, выделяющаяся при окислении неорганических соединений (хемоавтотрофы), у гетеротрофов — энергия окисления органических веществ (хемогетеротрофы).
Большинство живых организмов относится или к фотоавтотрофам (растения), или к хемогетеротрофам (грибы, животные). Если организмы, в зависимости от условий, ведут себя как авто- либо как гетеротрофы, то их называют миксотрофами (эвглена зеленая).
Репликация (редупликация) ДНК
Репликация ДНК — процесс самоудвоения, главное свойство молекулы ДНК. Репликация относится к категории реакций матричного синтеза, идет с участием ферментов. Под действием ферментов молекула ДНК раскручивается, и около каждой цепи, выступающей в роли матрицы, по принципам комплементарности и антипараллельности достраивается новая цепь. Таким образом, в каждой дочерней ДНК одна цепь является материнской, а вторая — вновь синтезированной. Такой способ синтеза называется полуконсервативным.
«Строительным материалом» и источником энергии для репликации являются дезоксирибонуклеозидтрифосфаты (АТФ, ТТФ, ГТФ, ЦТФ), содержащие три остатка фосфорной кислоты. При включении дезоксирибонуклеозидтрифосфатов в полинуклеотидную цепь два концевых остатка фосфорной кислоты отщепляются, и освободившаяся энергия используется на образование фосфодиэфирной связи между нуклеотидами.
В репликации участвуют следующие ферменты:
- геликазы («расплетают» ДНК);
- дестабилизирующие белки;
- ДНК-топоизомеразы (разрезают ДНК);
- ДНК-полимеразы (подбирают дезоксирибонуклеозидтрифосфаты и комплементарно присоединяют их к матричной цепи ДНК);
- РНК-праймазы (образуют РНК-затравки, праймеры);
- ДНК-лигазы (сшивают фрагменты ДНК).
С помощью геликаз в определенных участках ДНК расплетается, одноцепочечные участки ДНК связываются дестабилизирующими белками, образуется репликационная вилка. При расхождении 10 пар нуклеотидов (один виток спирали) молекула ДНК должна совершить полный оборот вокруг своей оси. Чтобы предотвратить это вращение ДНК-топоизомераза разрезает одну цепь ДНК, что дает ей возможность вращаться вокруг второй цепи.
ДНК-полимераза может присоединять нуклеотид только к 3′-углероду дезоксирибозы предыдущего нуклеотида, поэтому данный фермент способен передвигаться по матричной ДНК только в одном направлении: от 3′-конца к 5′-концу этой матричной ДНК. Так как в материнской ДНК цепи антипараллельны, то на ее разных цепях сборка дочерних полинуклеотидных цепей происходит по-разному и в противоположных направлениях. На цепи 3’–5′ синтез дочерней полинуклеотидной цепи идет без перерывов; эта дочерняя цепь будет называться лидирующей. На цепи 5’–3′ — прерывисто, фрагментами (фрагменты Оказаки), которые после завершения репликации ДНК-лигазами сшиваются в одну цепь; эта дочерняя цепь будет называться запаздывающей (отстающей).
Купить проверочные работы по биологии
Особенностью ДНК-полимеразы является то, что она может начинать свою работу только с «затравки» (праймера). Роль «затравок» выполняют короткие последовательности РНК, образуемые при участи фермента РНК-праймазы и спаренные с матричной ДНК. РНК-затравки после окончания сборки полинуклеотидных цепочек удаляются.
Репликация протекает сходно у прокариот и эукариот. Скорость синтеза ДНК у прокариот на порядок выше (1000 нуклеотидов в секунду), чем у эукариот (100 нуклеотидов в секунду). Репликация начинается одновременно в нескольких участках молекулы ДНК. Фрагмент ДНК от одной точки начала репликации до другой образует единицу репликации — репликон.
Репликация происходит перед делением клетки. Благодаря этой способности ДНК осуществляется передача наследственной информации от материнской клетки дочерним.
Некоторые свойства генетического кода
Генетический код практически универсален – за очень малым количеством исключений он одинаков у всех организмов, от бактерий до человека. Это свидетельствует, во-первых, о родстве всех форм жизни на Земле, а во-вторых, о древности самого кода. Вероятно, на ранних этапах существования примитивной жизни достаточно быстро сформировались разные варианты кода, но эволюционное преимущество получил только один.
Кроме того, он специфичен (однозначен): разные аминокислоты не кодируются одним и тем же триплетом. Также для генетического кода характерна вырожденность, или избыточность – несколько кодонов могут соответствовать одной и той же аминокислоте.
Считывание генетической записи осуществляется непрерывно; функции знаков препинания выполняют также триплеты оснований. Как правило, в генетическом «тексте» не бывает перекрывающихся записей, однако здесь тоже не обходится без исключений.
Этапы синтеза
Все процессы матричного синтеза поделены на три этапа:
- инициация (начало);
- элонгация;
- терминация (окончание).
Инициация представляет собой подготовку к синтезу, характер которой зависит от вида процесса. Главной целью этой стадии является приведение системы фермент-субстрат в рабочее состояние.
Во время элонгации непосредственно осуществляется наращивание синтезируемой цепи, при котором между подобранными согласно матричной последовательности звеньями замыкается ковалентная связь (пептидная или фосфодиэфирная). Терминация приводит к остановке синтеза и освобождению продукта.
Как происходит репликация
Матричный синтез ДНК происходит с участием множества ферментов и вспомогательных белков. Ключевыми компонентами являются:
- ДНК-хеликаза — расплетает двойную спираль, разрушает связи между цепями молекулы;
- ДНК-лигаза — «зашивает» разрывы между фрагментами Оказаки;
- праймаза — синтезирует затравку, необходимую для работы ДНК-синтезирующего фрагмента;
- SSB-белки — стабилизируют одноцепочечные фрагменты расплетенной ДНК;
- ДНК-полимеразы — синтезируют дочернюю матричную цепь.

Хеликаза, праймаза и SSB-белки подготавливают почву для синтеза. В результате каждая из цепей исходной молекулы становится матрицей. Синтез осуществляется с огромной скоростью (от 50 нуклеотидов в секунду).
Работа ДНК-полимеразы происходит в направлении от 5`к 3`- концу. Из-за этого на одной из цепей (лидирующей) синтез происходит по ходу расплетания и непрерывно, а на другой (отстающей) — в обратном направлении и отдельными фрагментами, названными «Оказаки».
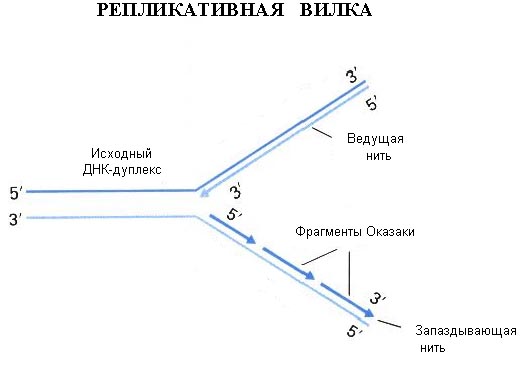
Y-образная структура, образованная в месте расплетания ДНК, называется репликационной вилкой.
Строение гена эукариот
Ген — участок молекулы ДНК, кодирующий первичную последовательность аминокислот в полипептиде или последовательность нуклеотидов в молекулах транспортных и рибосомных РНК. ДНК одной хромосомы может содержать несколько тысяч генов, которые располагаются в линейном порядке. Место гена в определенном участке хромосомы называется локусом. Особенностями строения гена эукариот являются: 1) наличие достаточно большого количества регуляторных блоков, 2) мозаичность (чередование кодирующих участков с некодирующими). Экзоны (Э) — участки гена, несущие информацию о строении полипептида. Интроны (И) — участки гена, не несущие информацию о строении полипептида. Число экзонов и интронов различных генов разное; экзоны чередуются с интронами, общая длина последних может превышать длину экзонов в два и более раз. Перед первым экзоном и после последнего экзона находятся нуклеотидные последовательности, называемые соответственно лидерной (ЛП) и трейлерной последовательностью (ТП). Лидерная и трейлерная последовательности, экзоны и интроны образуют единицу транскрипции. Промотор (П) — участок гена, к которому присоединяется фермент РНК-полимераза, представляет собой особое сочетание нуклеотидов. Перед единицей транскрипции, после нее, иногда в интронах находятся регуляторные элементы (РЭ), к которым относятся энхансеры и сайленсеры. Энхансеры ускоряют транскрипцию, сайленсеры тормозят ее.
Механизм транскрипции
Ключевым ферментом транскрипции является РНК-полимераза. Последняя бывает нескольких видов и отличается по строению у прокариот и эукариот. Однако механизм ее действия везде одинаков и заключается в наращивании цепи комплементарно подбираемых рибонуклеотидов с замыканием фосфодиэфирной связи между ними.
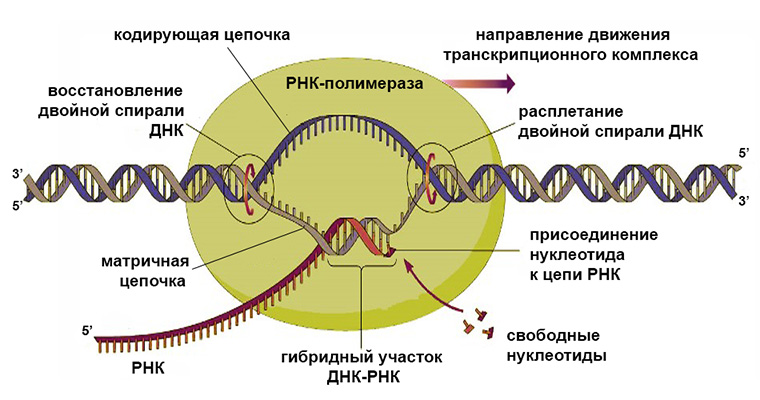
Матричной молекулой для этого процесса служит ДНК. На ее основе могут создаваться разные типы РНК, а не только информационные, которые используются в белковом синтезе.
Участок матрицы, с которого «списывается» последовательность РНК, называется транскриптоном. В его составе имеется промотор (место для присоединения РНК-полимеразы) и терминатор, на котором синтез останавливается.
Трансляция
Матричный синтез белка и у прокариот, и у эукариот осуществляется в специализированных органоидах — рибосомах. Последние состоят из двух субъединиц, одна из которых (малая) служит для связывания тРНК и матричной РНК, а другая (большая) принимает участие в образовании пептидных связей.
Началу трансляции предшествует активация аминокислот, т. е. присоединение их к соответствующим транспортным РНК с образованием макроэргической связи, за счет энергии которых впоследствии осуществляются реакции транспептидирования (присоединения к цепи очередного звена).
В процессе синтеза также принимают участие белковые факторы и ГТФ. Энергия последнего необходима для продвижения рибосомы по матричной цепи РНК.
Строение и функции РНК
РНК — полимер, мономерами которой являются рибонуклеотиды. В отличие от ДНК, РНК образована не двумя, а одной полинуклеотидной цепочкой (исключение — некоторые РНК-содержащие вирусы имеют двухцепочечную РНК). Нуклеотиды РНК способны образовывать водородные связи между собой. Цепи РНК значительно короче цепей ДНК.
Мономер РНК — нуклеотид (рибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания РНК также относятся к классам пиримидинов и пуринов.
Пиримидиновые основания РНК — урацил, цитозин, пуриновые основания — аденин и гуанин. Моносахарид нуклеотида РНК представлен рибозой.
Выделяют три вида РНК: 1) информационная (матричная) РНК — иРНК (мРНК), 2) транспортная РНК — тРНК, 3) рибосомная РНК — рРНК.
Все виды РНК представляют собой неразветвленные полинуклеотиды, имеют специфическую пространственную конформацию и принимают участие в процессах синтеза белка. Информация о строении всех видов РНК хранится в ДНК. Процесс синтеза РНК на матрице ДНК называется транскрипцией.
Транспортные РНК содержат обычно 76 (от 75 до 95) нуклеотидов; молекулярная масса — 25 000–30 000. На долю тРНК приходится около 10% от общего содержания РНК в клетке. Функции тРНК: 1) транспорт аминокислот к месту синтеза белка, к рибосомам, 2) трансляционный посредник. В клетке встречается около 40 видов тРНК, каждый из них имеет характерную только для него последовательность нуклеотидов. Однако у всех тРНК имеется несколько внутримолекулярных комплементарных участков, из-за которых тРНК приобретают конформацию, напоминающую по форме лист клевера. У любой тРНК есть петля для контакта с рибосомой (1), антикодоновая петля (2), петля для контакта с ферментом (3), акцепторный стебель (4), антикодон (5). Аминокислота присоединяется к 3′-концу акцепторного стебля. Антикодон — три нуклеотида, «опознающие» кодон иРНК. Следует подчеркнуть, что конкретная тРНК может транспортировать строго определенную аминокислоту, соответствующую ее антикодону. Специфичность соединения аминокислоты и тРНК достигается благодаря свойствам фермента аминоацил-тРНК-синтетаза.
Рибосомные РНК содержат 3000–5000 нуклеотидов; молекулярная масса — 1 000 000–1 500 000. На долю рРНК приходится 80–85% от общего содержания РНК в клетке. В комплексе с рибосомными белками рРНК образует рибосомы — органоиды, осуществляющие синтез белка. В эукариотических клетках синтез рРНК происходит в ядрышках. Функции рРНК: 1) необходимый структурный компонент рибосом и, таким образом, обеспечение функционирования рибосом; 2) обеспечение взаимодействия рибосомы и тРНК; 3) первоначальное связывание рибосомы и кодона-инициатора иРНК и определение рамки считывания, 4) формирование активного центра рибосомы.
Информационные РНК разнообразны по содержанию нуклеотидов и молекулярной массе (от 50 000 до 4 000 000). На долю иРНК приходится до 5% от общего содержания РНК в клетке. Функции иРНК: 1) перенос генетической информации от ДНК к рибосомам, 2) матрица для синтеза молекулы белка, 3) определение аминокислотной последовательности первичной структуры белковой молекулы.
Справочная информация
ДокументыЗаконыИзвещенияУтверждения документовДоговораЗапросы предложенийТехнические заданияПланы развитияДокументоведениеАналитикаМероприятияКонкурсыИтогиАдминистрации городовПриказыКонтрактыВыполнение работПротоколы рассмотрения заявокАукционыПроектыПротоколыБюджетные организацииМуниципалитетыРайоныОбразованияПрограммыОтчетыпо упоминаниямДокументная базаЦенные бумагиПоложенияФинансовые документыПостановленияРубрикатор по темамФинансыгорода Российской Федерациирегионыпо точным датамРегламентыТерминыНаучная терминологияФинансоваяЭкономическаяВремяДаты2015 год2016 годДокументы в финансовой сферев инвестиционной
Транскрипция и трансляция
Это два наиглавнейших шага биосинтеза.
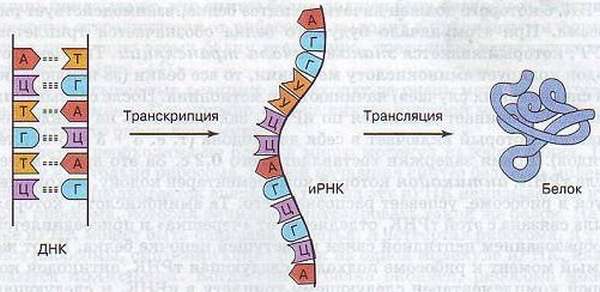
Транскрипция с латинского означает «переписывание» – в качестве матрицы применяется ДНК, поэтому происходит синтезирование трёх видов РНК (матричной/информационной, транспортной, рибосомной рибонуклеиновых кислот). Реакция осуществляется с помощью полимеразы (РНК) и с использованием большого количества аденозинтрифосфата.
Выделают два основных действия:
- Обозначение конца и начала трансляции присоединением иРНК.
- Событие, осуществляемое благодаря сплайсингу, который в свою очередь удаляет неинформационные последовательности РНК, тем самым происходит уменьшение массы матричной рибонуклеиновой кислоты в 10 раз.
Трансляция с латинского означает «перевод» – используется иРНК в качестве матрицы, синтезируются полипептидные цепочки.
Трансляция включает в себя три этапа, которые можно было представить в виде таблицы:
- Первый этап. Инициация формирование комплекса, который участвует в синтезе полипептидной цепочки.
- Второй этап. Элонгация увеличение размеров этой цепи.
- Третий этап. Терминация заключение выше упомянутого процесса.
Строение молекулы ДНК
Дезоксирибонуклеиновая кислота – это макромолекула, состоящая из двух нитей. Ее структура имеет несколько уровней организации.
Первичная структура цепи ДНК представляет собой последовательность нуклеотидов, содержащих каждый одно из четырех азотистых оснований: аденин, гуанин, цитозин или тимин. Цепочки возникают при соединении сахара дезоксирибозы одного нуклеотида с фосфатным остатком другого. Этот процесс осуществляется с участием белка-катализатора – ДНК-лигазы.
- Вторичная структура ДНК – это так называемая двойная спираль (точнее – двойной винт). Основания способны соединяться друг с другом следующим образом: аденин и тимин образуют двойную водородную связь, а гуанин и цитозин – тройную. Эта особенность лежит в основе принципа комплементарности оснований, согласно которому цепочки соединяются друг с другом. При этом происходит винтообразное (чаще — правое) закручивание двойной цепочки.
- Третичная структура – это сложная конформация огромной молекулы, возникающая посредством дополнительных водородных связей.
- Четвертичная структура образуется в комплексе с особыми белками и РНК и представляет собой способ упаковки ДНК в клеточном ядре.
Что представляют собой белки
Белок является важнейшим функциональным элементом в клетках всех живых организмов. Они выполняют каталитическую, структурную, регуляторную, сигнальную, защитную и множество других функций.
Молекула белка – это биополимер, образованный последовательностью аминокислотных остатков. Она, как и молекулы нуклеиновых кислот, характеризуется наличием нескольких уровней структурной организации – от первичного до четвертичного.
Существует 20 видов обладающих определенными особенностями (канонических) аминокислот, используемых живыми системами для построения огромного количества самых разнообразных белков. Самостоятельно белок, как правило, не синтезируется. Ведущая роль в формировании сложной белковой молекулы принадлежит нуклеиновым кислотам – ДНК и РНК.
Ферменты
В процессе репликации участвует большое количество различных ферментов. Каждый из них несет особую функцию в процессе.
ДНК – полимераза
ДНК – полимераза – это фермент, ответственный за создание ДНК из нуклеотидов, точнее строительных блоков для ДНК. ДНК – полимераза должна копировать молекулы двухцепочечной ДНК. Как раз именно этот процесс называется принципом полуконсервативности. Во время деления клетки ДНК занимается дублированием ее же самой. Так копия исходной молекулы передается к каждой из дочерних клеток вместе с генетической информацией.
Иногда же ДНК – полимераза может ошибиться, обычно это повторяется через каждый миллиард скопированных пар оснований. Поэтому данный фермент отвечает еще и за корректировку нити ДНК.
Также ДНК – полимераза может добавлять новые и свободные нуклеотиды к 3’ – концу, тем самым образуя новые нити. Однако ДНК – полимераза не может самостоятельно начинать новую цепь.
ДНК — лигаза
ДНК – лигаза – фермент, которые соединяет разрывы в отстающей цепочке ДНК. Лигаза образует фосфодиэфирные связи между свободными 3’ и 5’ концами. Для образования фосфодиэфирной связи ДНК – лигаза использует энергию, полученную из гидролиза (АТФ).
В 1961 году два американских генетика, а именно М. Мезельсон и Д. Вейгл, поняли, что при рекомбинации происходит разрыв и соединение ДНК. Это дало толчок ученым к поиску фермента, который мог бы сшивать разделенные фрагменты ДНК. В 1967 году М. Мезельсон и Д. Вейгл находят нужный фермент и называют его ДНК – лигазой.
ДНК – геликаза (хеликаза)
ДНК – геликаза – фермент, который раскручивает двухцепочечную спираль ДНК. Раскручивая цепи, данный фермент разделяет нити между собой, делая их одинарными. ДНК – геликаза движется по одноцепочечной нити, как только он встречает участок с двумя закрученными цепями, он разрывает водородные связи между основаниями, тем самым продвигая репликативную вилку дальше
ДНК – праймаза
Праймаза – фермент, необходимый для инициации репликации ДНК. Этот фермент синтезирует ДНК – праймеры, которые и запускают синтез матричной цепи. Также ДНК – праймеры запускают синтез фрагментов Оказаки для запаздывающей цепи.
ДНК – топоизомераза
ДНК – топоизомераза – фермент, который изменяет степень сверхспиральности2. Из-за суперспиральности образуется напряжение в спирали, которое в итоге мешает репликации. Чтобы такого не случалось, существует такой фермент как ДНК – топоизомераза.
Белки SSB
SSB – это белки, которые помогают сохранить нити ДНК в состоянии, когда они расплетены, а также они соединяют одноцепочечные фрагменты ДНК. Такие белки предотвращают комплементарное спаривание.
Строение и функции ДНК
ДНК — полимер, мономерами которой являются дезоксирибонуклеотиды. Модель пространственного строения молекулы ДНК в виде двойной спирали была предложена в 1953 г. Дж. Уотсоном и Ф. Криком (для построения этой модели они использовали работы М. Уилкинса, Р. Франклин, Э. Чаргаффа).
Молекула ДНК образована двумя полинуклеотидными цепями, спирально закрученными друг около друга и вместе вокруг воображаемой оси, т.е. представляет собой двойную спираль (исключение — некоторые ДНК-содержащие вирусы имеют одноцепочечную ДНК). Диаметр двойной спирали ДНК — 2 нм, расстояние между соседними нуклеотидами — 0,34 нм, на один оборот спирали приходится 10 пар нуклеотидов. Длина молекулы может достигать нескольких сантиметров. Молекулярный вес — десятки и сотни миллионов. Суммарная длина ДНК ядра клетки человека — около 2 м. В эукариотических клетках ДНК образует комплексы с белками и имеет специфическую пространственную конформацию.
Мономер ДНК — нуклеотид (дезоксирибонуклеотид) — состоит из остатков трех веществ: 1) азотистого основания, 2) пятиуглеродного моносахарида (пентозы) и 3) фосфорной кислоты. Азотистые основания нуклеиновых кислот относятся к классам пиримидинов и пуринов. Пиримидиновые основания ДНК (имеют в составе своей молекулы одно кольцо) — тимин, цитозин. Пуриновые основания (имеют два кольца) — аденин и гуанин.
Моносахарид нуклеотида ДНК представлен дезоксирибозой.
Название нуклеотида является производным от названия соответствующего основания. Нуклеотиды и азотистые основания обозначаются заглавными буквами.
| Азотистое основание | Название нуклеотида | Обозначение |
|---|---|---|
| Аденин | Адениловый | А (A) |
| Гуанин | Гуаниловый | Г (G) |
| Тимин | Тимидиловый | Т (T) |
| Цитозин | Цитидиловый | Ц (C) |
Полинуклеотидная цепь образуется в результате реакций конденсации нуклеотидов. При этом между 3′-углеродом остатка дезоксирибозы одного нуклеотида и остатком фосфорной кислоты другого возникает фосфоэфирная связь (относится к категории прочных ковалентных связей). Один конец полинуклеотидной цепи заканчивается 5′-углеродом (его называют 5′-концом), другой — 3′-углеродом (3′-концом).
Против одной цепи нуклеотидов располагается вторая цепь. Расположение нуклеотидов в этих двух цепях не случайное, а строго определенное: против аденина одной цепи в другой цепи всегда располагается тимин, а против гуанина — всегда цитозин, между аденином и тимином возникают две водородные связи, между гуанином и цитозином — три водородные связи. Закономерность, согласно которой нуклеотиды разных цепей ДНК строго упорядоченно располагаются (аденин — тимин, гуанин — цитозин) и избирательно соединяются друг с другом, называется принципом комплементарности. Следует отметить, что Дж. Уотсон и Ф. Крик пришли к пониманию принципа комплементарности после ознакомления с работами Э. Чаргаффа. Э. Чаргафф, изучив огромное количество образцов тканей и органов различных организмов, установил, что в любом фрагменте ДНК содержание остатков гуанина всегда точно соответствует содержанию цитозина, а аденина — тимину («правило Чаргаффа»), но объяснить этот факт он не смог.
Из принципа комплементарности следует, что последовательность нуклеотидов одной цепи определяет последовательность нуклеотидов другой.
Цепи ДНК антипараллельны (разнонаправлены), т.е. нуклеотиды разных цепей располагаются в противоположных направлениях, и, следовательно, напротив 3′-конца одной цепи находится 5′-конец другой. Молекулу ДНК иногда сравнивают с винтовой лестницей. «Перила» этой лестницы — сахарофосфатный остов (чередующиеся остатки дезоксирибозы и фосфорной кислоты); «ступени» — комплементарные азотистые основания.
Функция ДНК — хранение и передача наследственной информации.